

Dr. Rubén Gerardo Contreras Patiño
Investigador Cinvestav 3D
SNII Nivel II
- Licenciatura en Biología (1985). Facultad de Ciencias Biológicas, Universidad Autónoma del Estado de Morelos.
- Maestría en Ciencias (1998). Facultad de Medicina, UNAM.
- Doctorado en Ciencias (1991). Depto. de Fisiología, Biofísica y Neurociencias, CINVESTAV.
https://www.cinvestav.mx/zacatenco/fisiologia/conocimiento/directorio-de-investigacion/ruben-gerardo-contreras-patino
gerardo.contreras@cinvestav.mx
Teléfono
(+52) 55 5747 3800 +3369
Érika Yedid Del Oso Agustín
Asistente Secretarial
erika.deloso@cinvestav.mx
(+52) 55 5747 3800 +5182
erika.deloso@cinvestav.mx
(+52) 55 5747 3800 +5182
Línea de Investigación:
Nuestro laboratorio se dedica a descifrar cómo las células se comunican y organizan a través de sus uniones intercelulares. Exploramos estos mecanismos fundamentales en condiciones fisiológicas y en enfermedades, con un enfoque en tres áreas clave:
• Riñón e Hipertensión: Cómo el alto consumo de sal afecta a los epitelios renales.
• Cáncer Cervicouterino: El papel de las uniones celulares en modelos de cáncer cervical.
• Neurociencia y Alzheimer: Estudiamos la barrera endotelial cerebral en modelos de esta enfermedad.
Enfoque Metodológico y Tecnologías
Combinamos modelos experimentales (ratas hipertensas, ratones transgénicos) con técnicas de vanguardia para obtener una visión integral:
• Cultivo celular y manipulación génica.
• Evaluación funcional (resistencia eléctrica transepitelial).
• Análisis molecular (inmunoblot, genes reporteros, secuenciación de tercera generación).
• Visualización avanzada (microscopía confocal e inmunofluorescencia).
Nuestro laboratorio se dedica a descifrar cómo las células se comunican y organizan a través de sus uniones intercelulares. Exploramos estos mecanismos fundamentales en condiciones fisiológicas y en enfermedades, con un enfoque en tres áreas clave:
• Riñón e Hipertensión: Cómo el alto consumo de sal afecta a los epitelios renales.
• Cáncer Cervicouterino: El papel de las uniones celulares en modelos de cáncer cervical.
• Neurociencia y Alzheimer: Estudiamos la barrera endotelial cerebral en modelos de esta enfermedad.
Enfoque Metodológico y Tecnologías
Combinamos modelos experimentales (ratas hipertensas, ratones transgénicos) con técnicas de vanguardia para obtener una visión integral:
• Cultivo celular y manipulación génica.
• Evaluación funcional (resistencia eléctrica transepitelial).
• Análisis molecular (inmunoblot, genes reporteros, secuenciación de tercera generación).
• Visualización avanzada (microscopía confocal e inmunofluorescencia).

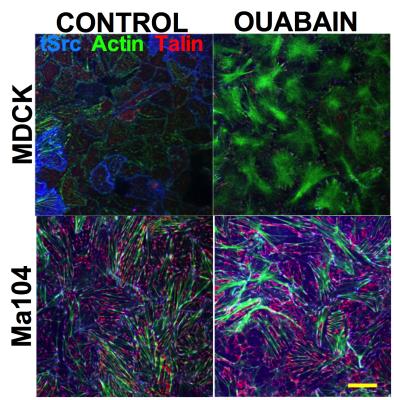
Ponce A, Flores-Maldonado C, Rubén CG.
Biomolecules. 2025; 16(6): 885. doi: 10.3390/biom15060885.
Biomolecules. 2025; 16(6): 885. doi: 10.3390/biom15060885.
Contreras RG, Torres-Carrillo A, Flores-Maldonado C, Shoshani L, Ponce A.
Int J Mol Sci. 2024; 25(11). doi: 10.3390/ijms25116122.
Int J Mol Sci. 2024; 25(11). doi: 10.3390/ijms25116122.
Campos-Blázquez JP, Flores-Maldonado C, Gallardo JM, Bonilla-Delgado J, Pedraza-Ramírez AA, López-Méndez O, Cortés-Malagón EM, Contreras RG.
Autophagy Rep. 2023; 2(1):2256146. doi: 10.1080/27694127.2023.2256146.
Autophagy Rep. 2023; 2(1):2256146. doi: 10.1080/27694127.2023.2256146.
Campos-Blázquez JP, Schuth N, Garay E, Clark AH, Vogelsang U, Nachtegaal M, Contreras RG, Quintanar L, Missirlis F.
Metallomics. 2022; 14(10). doi: 10.1093/mtomcs/mfac075
Metallomics. 2022; 14(10). doi: 10.1093/mtomcs/mfac075
Estrada N, Núñez-Vázquez EJ, Palacios A, Ascencio F, Guzmán-Villanueva L, Contreras RG.
Front Immunol. 2021; 12: 634497. doi: 10.3389/fimmu.2021.634497.
Front Immunol. 2021; 12: 634497. doi: 10.3389/fimmu.2021.634497.
Castillo A, Ortuño-Pineda C, Flores-Maldonado C, Larre I, Martínez Rendón J, Hinojosa L, Ponce A, Ogazón A, Serrano M, Valdes J, Contreras RG, Cereijido M.
Cell Physiol Biochem. 2019; 52(6): 1381-1397. doi: 10.33594/000000097.
Cell Physiol Biochem. 2019; 52(6): 1381-1397. doi: 10.33594/000000097.
Bonilla-Delgado J, Bulut G, Liu X, Cortés-Malagón EM, Schlegel R, Flores-Maldonado C, Contreras RG, Chung SH, Lambert PF, Uren A, Gariglio P.
Mol Cancer Res. 2012; 10(2): 250-8. doi: 10.1158/1541-7786.MCR-11-0287.
Mol Cancer Res. 2012; 10(2): 250-8. doi: 10.1158/1541-7786.MCR-11-0287.
Estudiantes de Doctorado
Jessica Paulina Campos Blázquez
jessica.campos@cinvestav.mx
Alan Arturo Pedraza Ramírez
alan.pedraza@cinvestav.mx
Octavio López Méndez
octavio.lopez@cinvestav.mx
Personal
Catalina Elizabeth Flores Maldonado
Auxiliar de Investigación
catalina.flores@cinvestav.mx
catalina.flores@cinvestav.mx











